インターフェロン-ガンマ依存的な抗トキソプラズマ応答におけるオートファジー蛋白質の役割
Role of Mouse and Human Autophagy Proteins in IFN-γ-Induced Cell-Autonomous Responses against Toxoplasma gondii.
Journal of Immunology Available online 21 Feb 2014 論文
概要
オートファジー必須分子群の一部が、インターフェロン-ガンマ依存的なトキソプラズマ原虫「殺傷」機構でとても重要な役割を果たしていることを発見しました。
背景
インターフェロン-ガンマ(IFN-γ)は細胞内寄生性原虫であるトキソプラズマに対する細胞内自然免疫応答を引き起こします。その応答に重要な役割を果たすIFN-γ誘導性GTP分解酵素であるIRGやGBPは、バクテリアに対してオートファジーによる応答を生じることが知られています。また、オートファジー必須分子であるAtg5は以前に抗トキソプラズマ応答に重要であることが報告されていますが、他のオートファジー必須蛋白質の役割については不明のままでした(図1)。
研究の成果
我々はAtg7、Atg9、Atg14LやAtg16L1といったオートファジー蛋白質を欠損するマウス胎児由来線維芽細胞(MEF)を用いて、抗トキソプラズマ応答について検討しました。Atg7やAtg16L1を欠損する細胞では、IFN-γ刺激による原虫数の低下が著しく阻害されました。また、IRGやGBPの原虫周囲への蓄積もAtg7やAtg16L1を欠損する細胞では低下していました(図2、図3)。
一方、Atg9やAtg14Lを欠損する細胞においては、IFN-γ刺激により野生型細胞と同程度に原虫が少なくなっており、IRGやGBPの原虫周辺への蓄積も認められました(図4、図5)。
以上のことから、マウスにおけるIFN-γ依存的な抗トキソプラズマ応答はオートファジー蛋白質Atg5に加えて、Atg7/Atg16L1も重要な役割を果たしていること、さらにAtg9やAtg14には非依存的であることが明らかになりました(図6)。
これらの結果から、IFN-γ依存的なトキソプラズマ原虫殺傷機構は、完全な“オートファジー”とはいえず、むしろオートファジーでも利用されているAtg7/Atg16L1 依存的な「膜輸送(メンブレントラフィッキング)」が、IFN-γ依存的殺傷機構にも使用されていると考えられます。
また、米国Duke大学のJorn Coer博士との共同研究により、Atg8の脂質付加反応に重要かつオートファジー必須分子であるAtg3も、IFN-γ依存的原虫殺傷機構に重要であることを示しました(研究業績の文献98参照。リンクはこちら)。今後は、Atg3/5/7/16L1がどのようにしてIRGやGBPによるトキソプラズマ原虫寄生胞膜破壊システムを制御しているのかを明らかにしていく予定です。
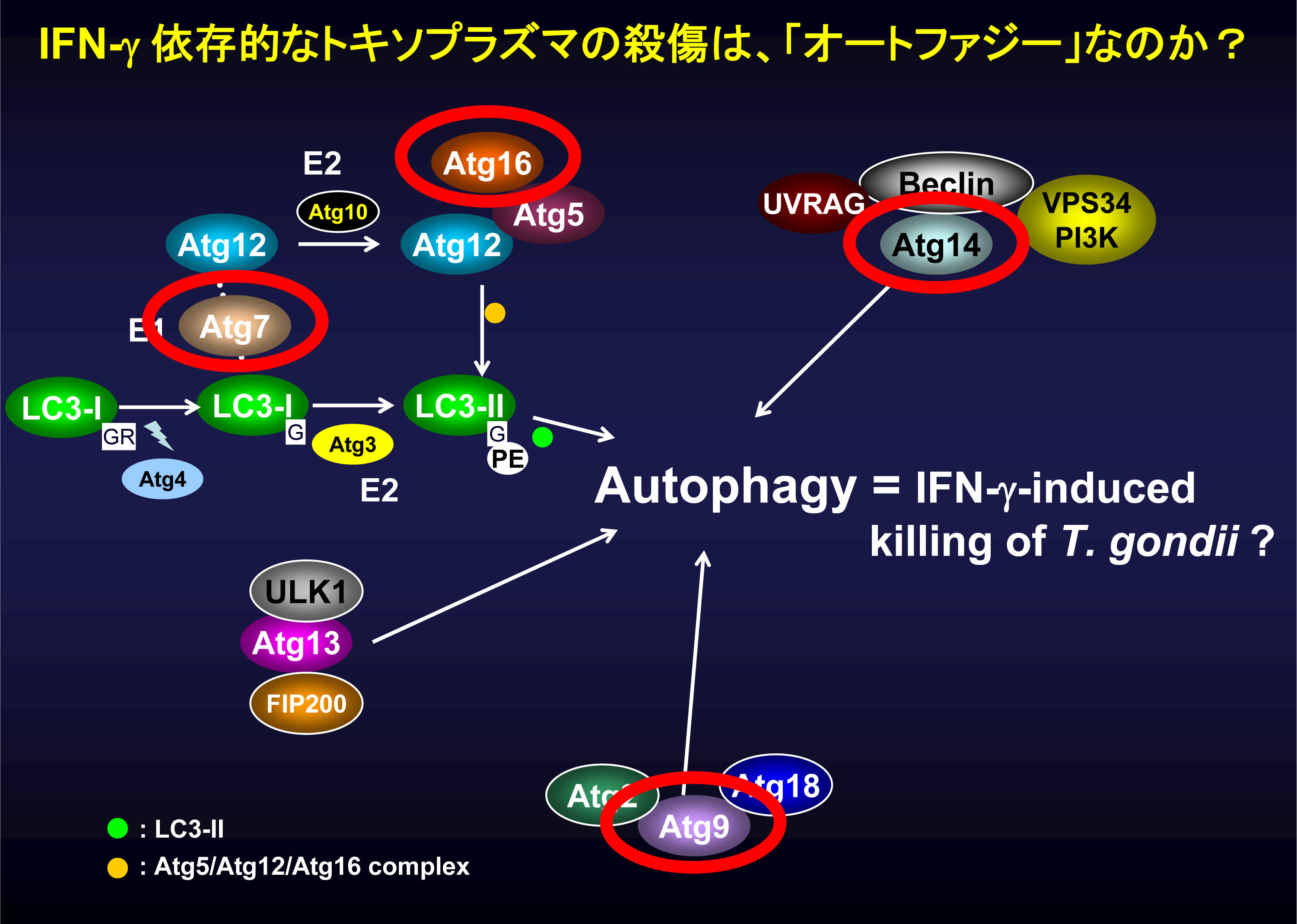
図1 オートファジー必須分子群の概略図
オートファジー必須分子群のうち、Atg5はIFN-γ依存的な抗トキソプラズマ応答に重要であることが報告されているが、他の分子の関与についてはまだ不明のままである。そこで我々は、オートファジー必須分子であるAtg7、Atg16L1、Atg9、Atg14Lについて、IFN-γ依存的原虫殺傷機構への関わりを検討した。
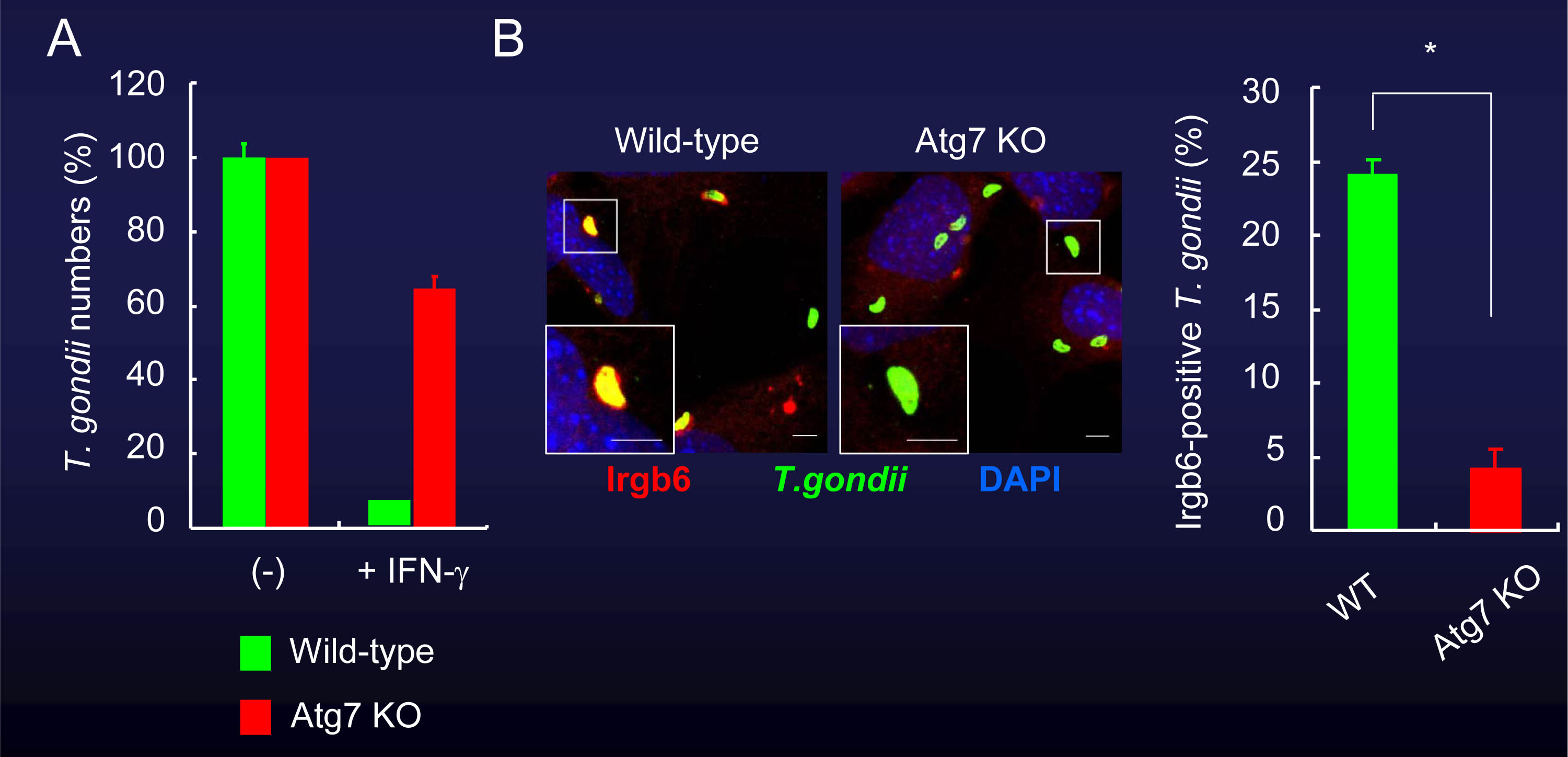
図2 Atg7欠損細胞では、IFN-γ刺激による細胞内のトキソプラズマ原虫への殺傷能が減弱する。
(A)Atg7を欠いた細胞では、IFN-γ刺激時の原虫数の低下が著しく阻害される。
(B)Atg7を欠いた細胞では、IRGの原虫周囲への蓄積も減少していた。
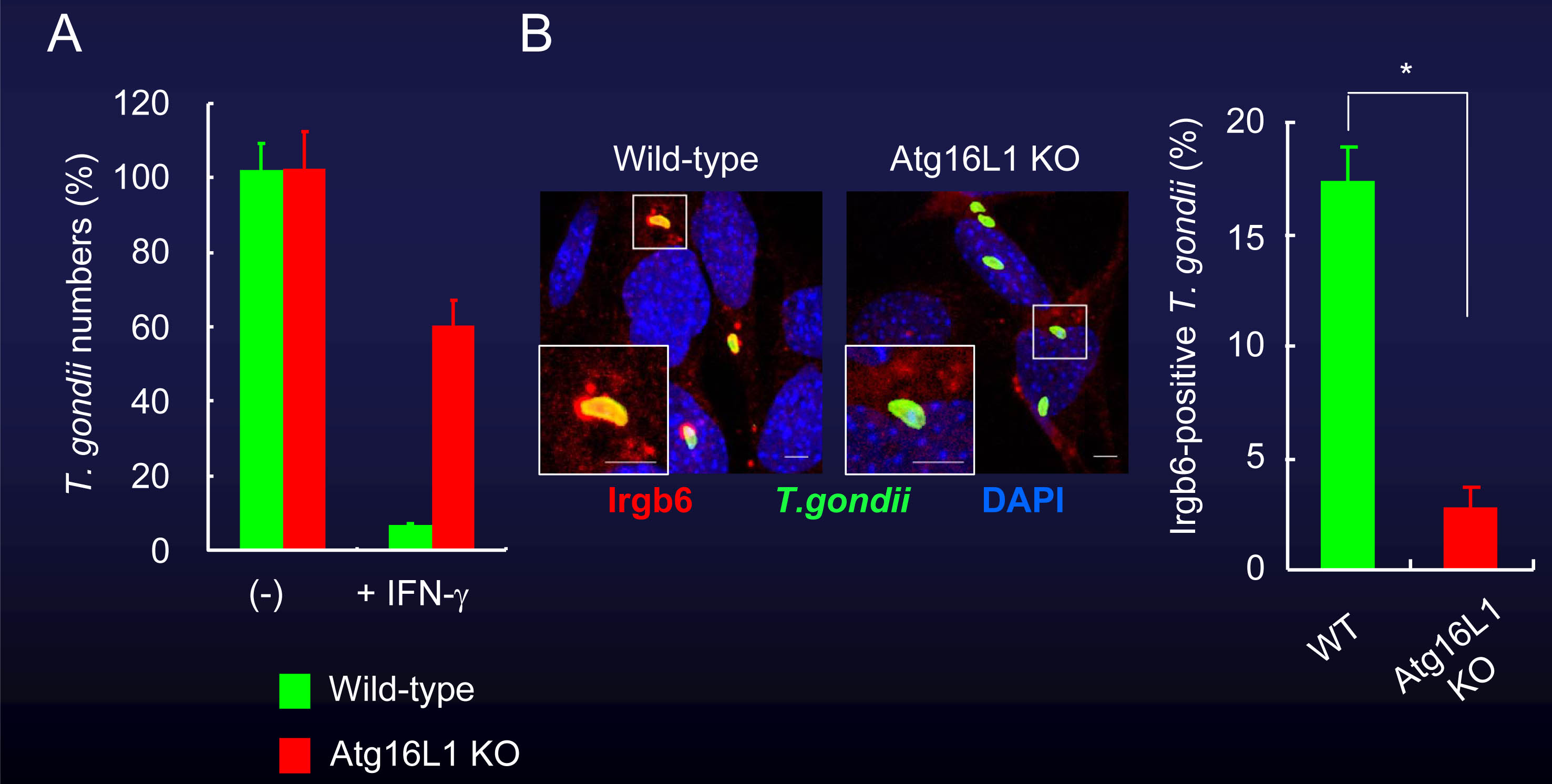
図3 Atg16L1欠損細胞でも、IFN-γ刺激による細胞内のトキソプラズマ原虫への殺傷能が減弱する。
(A)Atg16L1を欠いた細胞では、IFN-γ刺激時の原虫数の低下が著しく阻害される。
(B)また、Atg16L1を欠いた細胞では、IRGの原虫周囲への蓄積も減少していた。
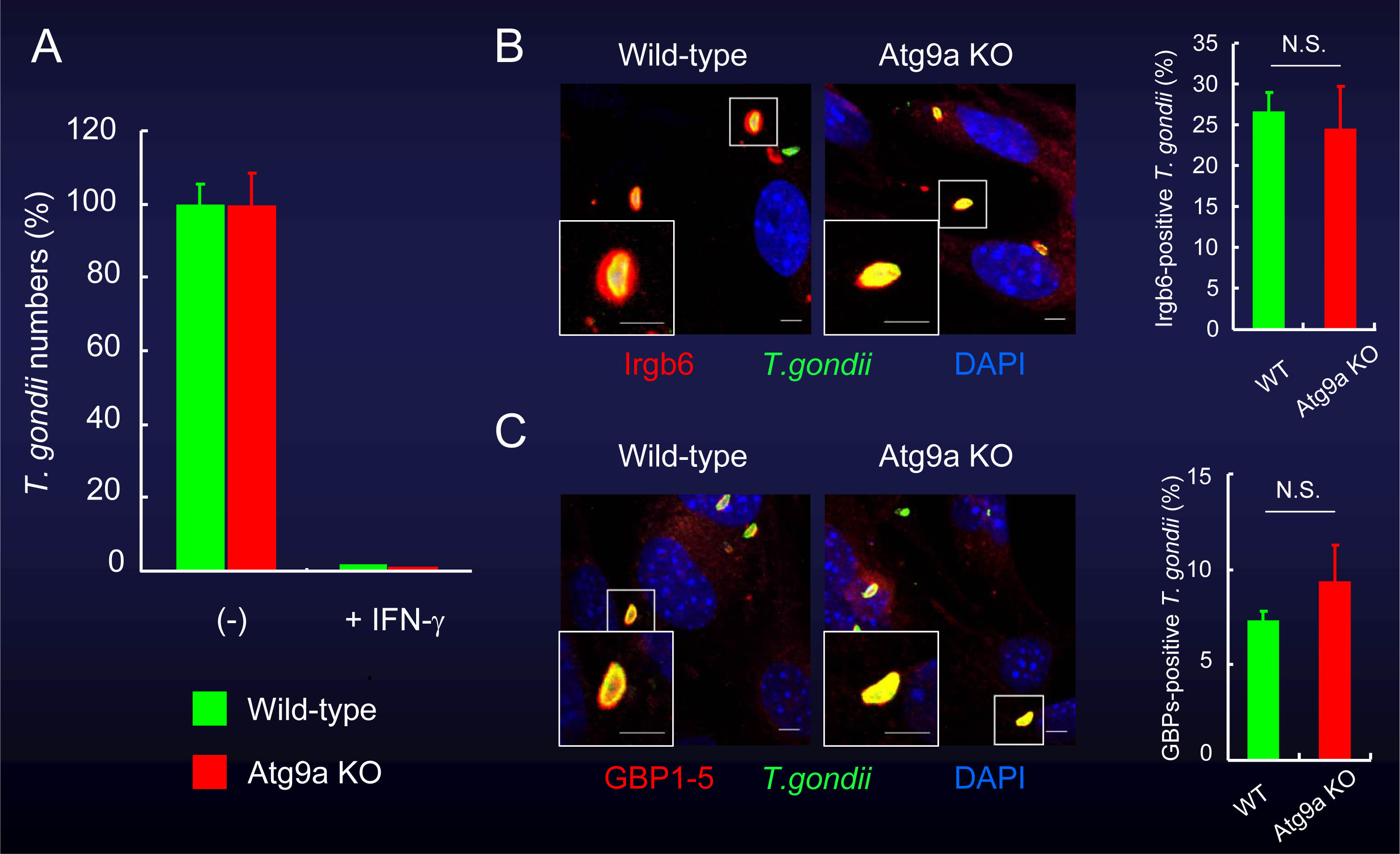
図4 Atg9欠損細胞では、IFN-γ刺激による細胞内のトキソプラズマ原虫への殺傷能は野生型細胞と同程度に見られる。
(A)Atg9を欠損しても、IFN-γ刺激時の原虫数の低下には影響しない。
(B)(C)また、Atg9欠損細胞では、IRGやGBPの原虫周囲への集積が野生型細胞と同程度に認められた。
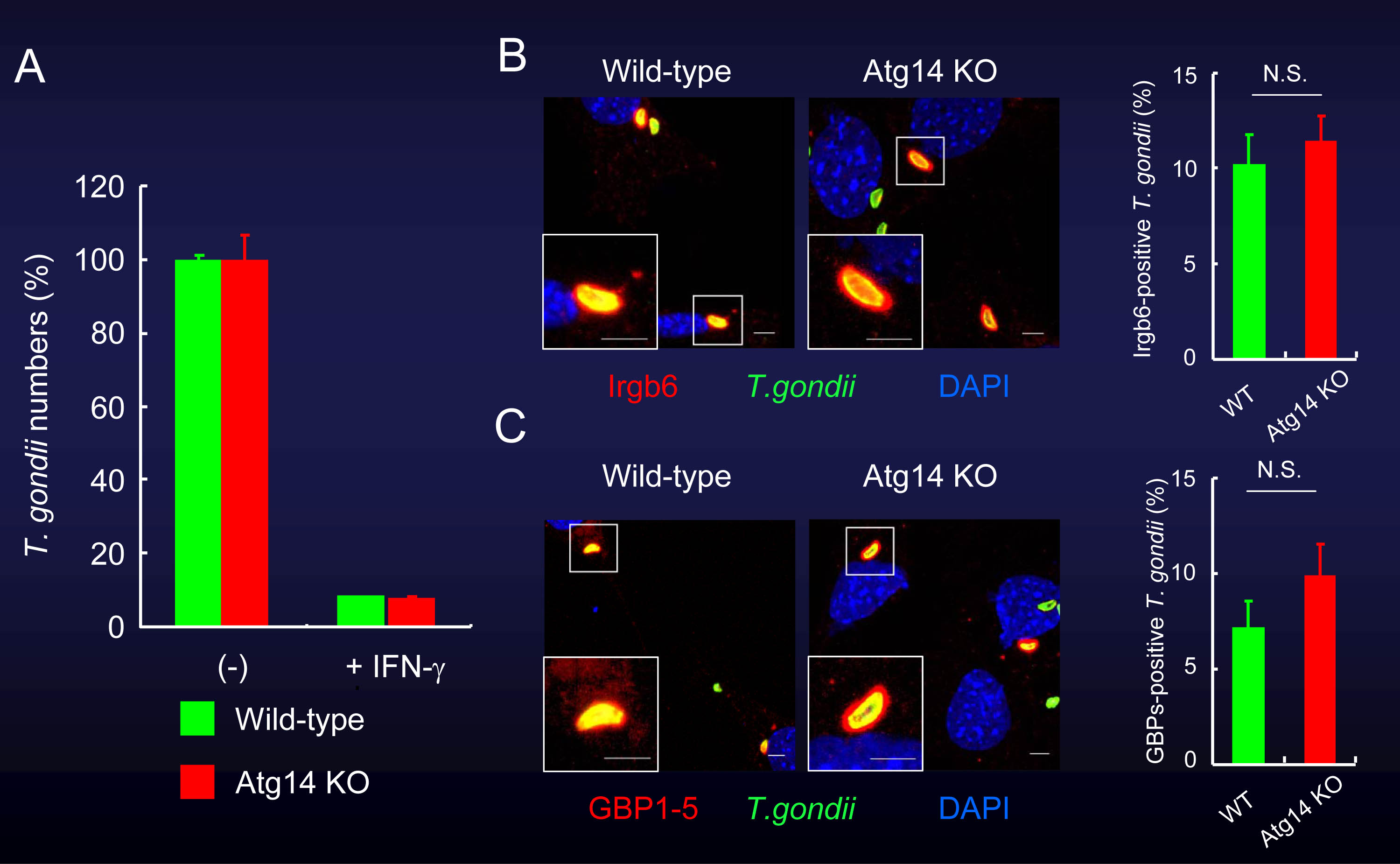
図5 Atg14L欠損細胞でも、IFN-γ刺激による細胞内のトキソプラズマ原虫への殺傷能は野生型細胞と同程度に見られる。
(A)Atg14Lを欠損しても、IFN-γ刺激時の原虫数の低下には影響しない。
(B)(C)また、Atg14L欠損細胞では、IRGやGBPの原虫周囲への集積が野生型細胞と同程度に認められた。
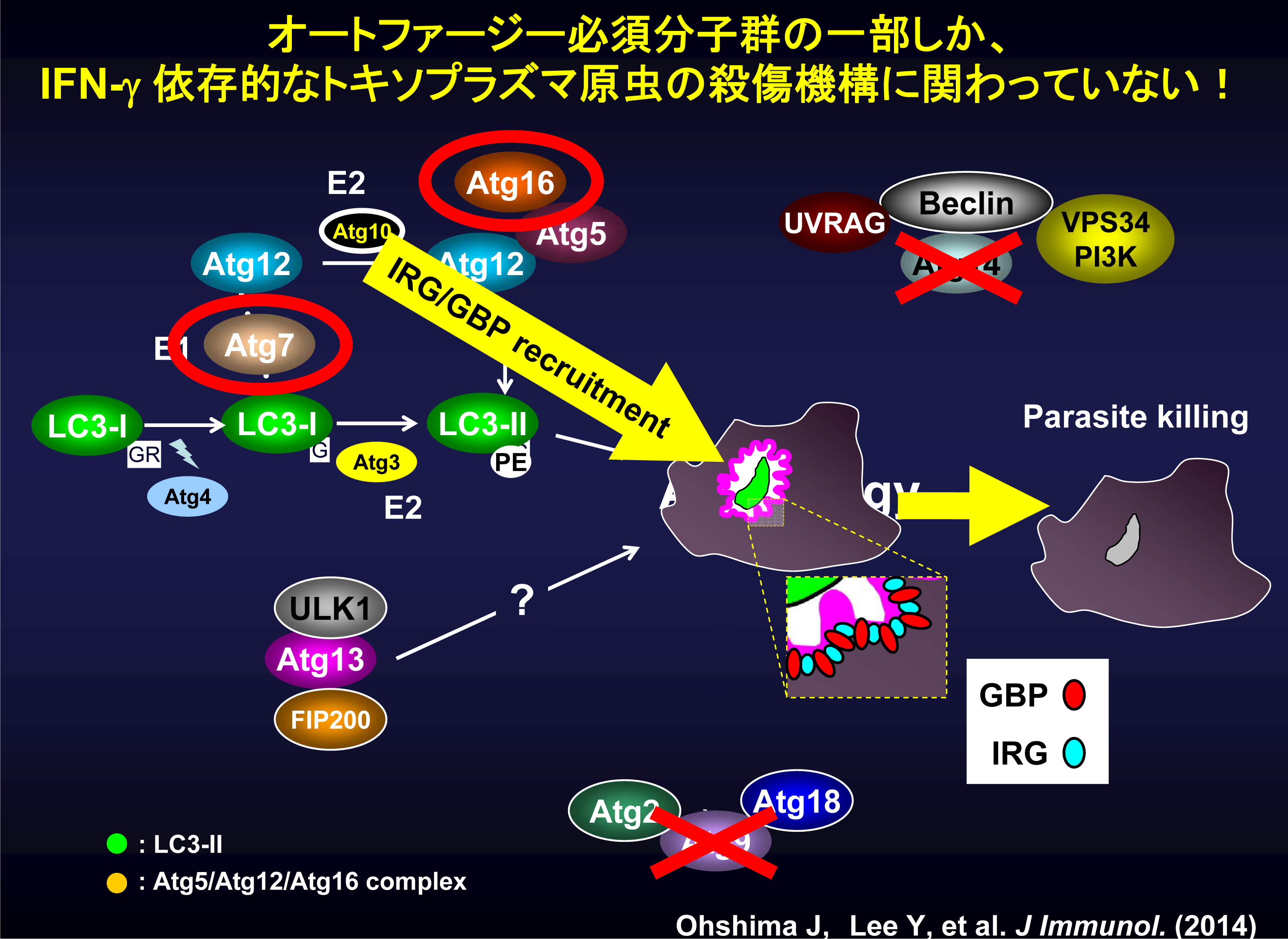
図6 IFN-γ依存的なトキソプラズマ原虫殺傷機構には、オートファジー必須分子群のうちの一部だけが 関与している。
-
論文詳細12
詳細を見る -
論文詳細11
詳細を見る -
論文詳細10
詳細を見る -
論文詳細09
詳細を見る -
論文詳細08
ヒトの細胞で抗トキソプラズ免疫反応に重要な分子の同定
―宿主分子IDO1を中心とした宿主-病原体相互作用の解明―
Toxoplasma Effector TgIST Targets Host IDO1 to Antagonize the IFN-γ-Induced Anti-parasitic Response in Human Cells
詳細を見る -
論文詳細07
抗病原体分子をバランスよく配置して、免疫反応を効率化!
―GBP依存的な抗病原体免疫反応に重要な制御因子Gate-16の同定―
Essential role for GABARAP autophagy proteins in interferon-inducible GTPase-mediated host defense
詳細を見る -
論文詳細06
実験的トキソプラズマワクチンがどのように効くかを解明!
―宿主タンパク質 p62を標的とした新たなワクチンの開発に前進―
p62 Plays a Specific Role in Interferon-γ-Induced Presentation of a Toxoplasma Vacuolar Antigen
詳細を見る -
論文詳細05
世界初!寄生虫「トキソプラズマ」に対する免疫反応が大幅アップ!
―抗トキソプラズマ免疫反応のブレーキ役を果たす分子の同定―
RabGDIα is a negative regulator of interferon-γ-inducible GTPase-dependent cell-autonomous immunity to Toxoplasma gondii
詳細を見る -
論文詳細04
寄生虫「トキソプラズマ」は、どのように宿主の身体を乗っ取るか?
―高病原性因子 高病原性因子 GRA6「トロイの木馬」現象を引き起こす ―
Selective and strain-specific NFAT4 activation by the Toxoplasma gondii polymorphic dense granule protein GRA6.
詳細を見る -
論文詳細03
インターフェロン-ガンマ依存的な抗トキソプラズマ応答におけるオートファジー蛋白質の役割
Role of Mouse and Human Autophagy Proteins in IFN-γ-Induced Cell-Autonomous Responses against Toxoplasma gondii.
詳細を見る -
論文詳細02
CREBHによるスルピリンの副作用誘導機構
CREBHはスルピリンによって誘導される致死的ショックの重症度を決定する
CREBH Determines the Severity of Sulpyrine-induced Fatal Shock
詳細を見る -
論文詳細01
トキソプラズマ症の発病を防ぐ最重要宿主防御因子GBPの同定
インターフェロン誘導性タンパクGBPは寄生虫「トキソプラズマ」を破壊する
A Cluster of Interferon-γ-Inducible p65 GTPases Plays a Critical Role in Host Defense against Toxoplasma gondii
詳細を見る